

 +34Kontinente:EUAS
+34Kontinente:EUAS







































1. Lebendfotos
1.1. Falter
1.1.1. bisexuelle Form



1.1.2. parthenogenetische Form(en)













1.2. Raupe (parthenogenetische Form(en)














1.3. Puppe (parthenogenetische Form(en)



1.4. Ei (parthenogenetische Form(en)
2. Diagnose
2.1. Männchen (bisexuelle Form)

2.2. Raupe
"Typisch für Dahlica triquetrella sind der braune Kopf, der breite Sack und die angesponnenen Chitinteile (letzteres auch bei Diplodoma, aber der Sack sieht anders aus)." [Erwin Hauser]
2.3. Genitalien
2.3.1. Weibchen
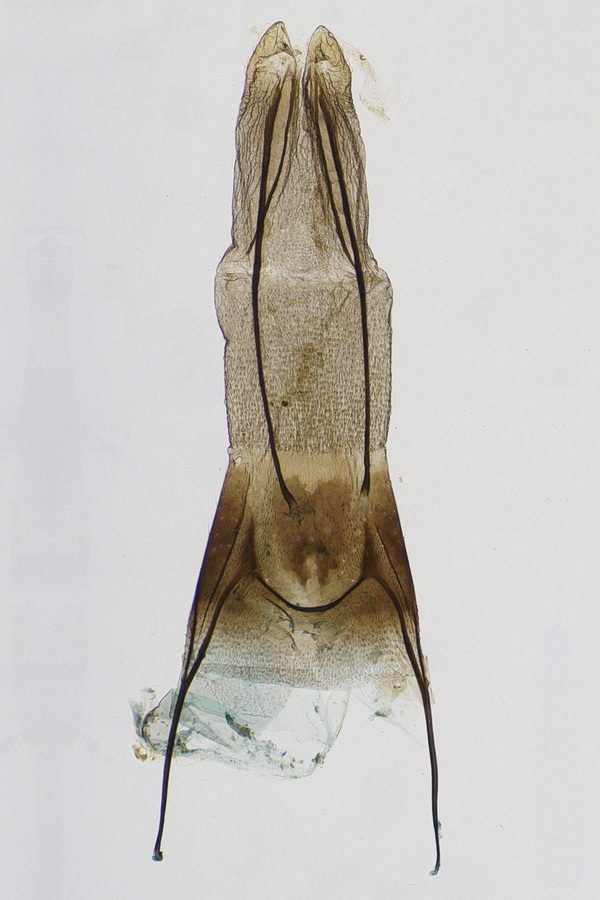
2.4. Erstbeschreibung

3. Biologie
3.1. Habitat (bisexuelle Form)


3.2. Habitat (parthenogenetische Form(en))



4. Weitere Informationen
4.1. Etymologie (Namenserklärung)
„triquetrus dreieckig, nach der Sackform.“
Sauter (1954) begründet seine Namenswahl seiner Solenobia seileri (jetzt Unterart): "Ich widme diese ihrer Parthenogenese wegen besonders interessante neue Art meinem verehrten Lehrer, Herrn Prof. Dr. J. Seiler. Gleichzeitig möchte ich ihm herzlich danken für die Überlassung des Materials und für die stetige Unterstützung meiner Arbeit."
4.2. Andere Kombinationen
- Tinea triquetrella Hübner, 1813 [Originalkombination]
4.3. Synonyme
- Dahlica larviformis Enderlein, 1912
- Solenobia thurneri Sieder, 1953
- 00763 Solenobia seileri Sauter, 1954 [jetzt Subspecies]
- 00763 Dahlica seileri (Sauter, 1954) [jetzt Subspecies]
4.4. Nomenklatur und Taxonomie
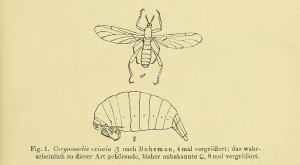
Die Gattung Dahlica sowie die Species D. larviformis wurden von Enderlein (1912: 264) für ein Tier aufgestellt, das er für eine Fliege hielt; er stellte das Taxon vorläufig zu den Mycetophiliden. Die Originalbeschreibung von D. larviformis besteht aus wenigen Zeilen; ausführlicher ist eine Arbeit des patronymisch Geehrten Prof. Dr. Dahl (1911), aus der hier der erste Absatz zitiert sei: „Am 10. Mai 1908 fand ich in der Nähe von Berlin (Bei Hermsdorf) an einem Kiefernstamm, etwa 1¼ m über dem Boden, auf einer leeren Schmetterlingspuppe ein eigenartiges Tier. Ich hielt dasselbe zuerst für irgend eine Mißgeburt und steckte es, ohne die Lebensbedingungen näher zu prüfen, in Spiritus. – Zu Hause erwies sich sich als ein höchst eigenartiges Insekt, als ein flügel- und schwingerloser Zweiflügler (Fig. 1 ♀).“

(Autor: Jürgen Rodeland)
4.5. Taxonomie und Faunistik
Vorabbemerkung: Arnscheid & Weidlich (2017: 39) betonen bei der Einführung der Gattung Dahlica rückblickend, dass alle bisher zur Arttrennung genutzten Merkmale (Genital-Indices, Schuppen-Klassen-Indices, Flügelgeäder) eine deutlich größere intraspezifische Variation aufweisen, als zunächst gedacht. Dies machte die Artabgrenzung schwierig und führte zu einer Unzahl widersprüchlicher Artbeschreibungen und Fundortmeldungen. Mittlerweile wurden auch genetische Aspekte - insbesondere durch Barcoding, teilweise auch durch die Untersuchung von Kern-DNA - mit einbezogen. Dies verhalf teilweise zur Klärung, warf aber zahlreiche neue Fragen auf. Arnscheid & Weidlich (2017: 39) kündigen daher an, dass wir auch in der nahen Zukunft mit einem öfteren Durcheinanderwirbeln der Taxa rechnen müssen: "It is foreseeable that further studies of molecular characteristics will substantially change the taxonomy of the tribe in the near future (Chevasco et al. 2014, Elzinga et al. 2014)."
Dahlica triquetrella agg. gehört zu den noch relativ klar ansprechbaren Arten, zumindest dann, wenn man bisexuelle und parthenogenetische Formen bei einer Art zusammenlässt.
Wir hatten hier zunächst für die bisexuelle und die parthenogenetische Form getrennte Seiten geführt. Doch die Sache ist noch komplizierter, da es mindestens 2 parthenogenetische Formen gibt, eine diploide und eine tetraploide. Und angesichts der Barcoding-Ergebnisse scheinen die parthenogenetischen Formen noch nicht wirklich von der sich sexuell fortpflanzenden Form entfernt zu sein. Wir fassen - wie auch Arnscheid & Weidlich (2017: 42-43) - wieder alles unter einem Artnamen zusammen: Dahlica triquetrella.
Weidlich (2014) erstellt einen Aufsatz "Zum Vorkommen der bisexuellen Form von Dahlica triquetrella (Hübner, 1813) in Europa, insbesondere in Ungarn und in der Slowakei". Er betont dabei die insgesamt große, aber meist nur punktuelle Verbreitung der Art in vielen Teilen des Vorkommens. In Deutschland ist die bisexuelle Form der Art nach bisheriger Kenntnis auf Baden-Württemberg (Adelegg) und Bayern beschränkt; die Angabe aus Sachsen ist unsicher.
Diese Form mit regulär zwei Geschlechtern gilt als Glazialrelikt, das weitgehend (oder vollständig?) an die Alpen gebunden ist. Arnscheid & Weidlich (2017: 42-43) fassen zur Verbreitung dieser Form zusammen: "The bisexual form is distributed very locally in scattered colonies and rather scarce in Germany, Switzerland and Austria. Records from the Netherlands, Belgium, Luxembourg, Czech Republic and Hungary (Sauter & Hättenschwiler, 1996: 39, Arnscheid, 2004) need further confirmation and are probably due to misinterpretation of recorded larval cases belonging to the parthenogenetic form." Auch aus Italien gibt es keinen sicheren Nachweis dieser Form.
Parthenogenetische Formen gibt es auch in den Alpen. Insgesamt ist die diploid parthenogenetische Form aber sehr, sehr viel weiter verbreitet. Die Vorkommen reichen zum einen südwärts bis einschließlich Spanien, Italien, Balkan (aber ohne Portugal und ohne die meisten Inseln), zum anderen nordwärts quer durch Frankreich und Deutschland bis Skandinavien und ostwärts nach Sibirien und Ost-Asien. Arnscheid & Weidlich (2017: 42) tragen neuere Funde aus Kroatien, Ungarn, Slowenien und der Slowakei zusammen.
Weidlich (2015: 1915) stellt eigene Fundnachweise der parthenogenetischen Form für Italien zusammen: "Fundnachweise: 1995: Monte Simeone; 1995: Ronchi; 1995, 1999: Iamiano; 1996: Roccaraso; 1997: Colle Melosa; 2003: Opi; 2003: Prati di Tivo; 2005: Pordoijoch; 2015: Monte Arazecca." Er erläutert: "Nur relativ wenige Nachweise, stellenweise aber auch häufig und südlich bis Opi im Nationalpark Abruzzen; zwischen 40 und 2.500 m NN nachgewiesen."
Das Taxon Dahlica seileri (Sauter, 1954) erlebte eine sehr wechselvolle Geschichte: Es wurde aus der Schweiz beschrieben, aber seit Längerem in schweizerischen Arbeiten meist nicht mehr akzeptiert (und oft genug auch gar nicht mehr erwähnt)! Bei Karsholt & Razowski (1996), der Fauna Europaea und im World Catalogue von Sobczyk (2011) wurde das Taxon noch als Art geführt.
Sauter (1954) formulierte in seiner Erstbeschreibung: "Diese der triquetrella sehr nahestehende Art ist bisher nur in der parthenogenetischen Form bekannt. Sie wurde erstmals von Goppenstein zusammen mit triquetrella eingetragen. Die Säcke sind aber von denen von triquetrella etwas verschieden, was sich noch deutlicher zeigte, als die Art gezüchtet wurde (genaue Angaben über den Sack wird die oben erwähnte Arbeit über die Präimaginalstadien bringen). Aus einer Kultur Seilers dieser parthenogenetischen Form schlüpfte nun 1954 ein einzelnes Männchen. Dieses weicht ebenfalls in einigen Punkten von triquetrella ab."
Es folgt die Beschreibung der beiden Geschlechter, die - wegen dem lange umstrittenen Status und der daraus abzuleitenden weiteren Diskussion - hier vollständig wiedergegeben wird:
"♂: Gross, breitflügelig, Flügelform wie triquetrella. Weisse Flecke der Vorderflügel klein und scharf (also ähnlich wie bei manni oder pineti, während sie bei triquetrella meistens grösser sind). Am Ende der Discoidalzelle ein grosser, dunkler Fleck vorhanden. Deckschuppen erheblich breiter als bei triquetrella, 4—6zackig, mindestens so breit wie bei den breitschuppigsten mir vorliegenden pineti-Tieren. Fransen grau, kaum heller als die Flügelgrundfarbe. Vorderflügellänge 8,3 mm. Vordertibie ohne Sporn (d. h. wie triquetrella, während sie bei pineti einen Subapicalsporn besitzt).
♀: Sehr ähnlich triquetrella, doch sind selten alle Tarsen rein 5-gliedrig, meistens treten Verschmelzungen auf, es wurden auch Tiere mit einzelnen rein 4-gliedrigen Tarsen beobachtet (bei triquetrella meist 5-gliedrig, 4-gliedrige Tarsen habe ich hier nicht gefunden). Ähnliche Tarsenbildung zeigt fumosella, deren ♀ aber sehr schmale Deckschuppen aufweist und deshalb nicht mit seileri verwechselt werden kann.
Diese Art wurde an Felsblöcken bei Goppenstein (Wallis) gefunden. Sie scheint dort viel seltener zu sein als triquetrella."
In der Folge wurde das Taxon vielfach als Form von Dahlica triquetrella angesehen, und auch der Erstbeschreiber bekam Bedenken bezüglich des Artstatus. Nach eigener Aussage hatte er die Art vor allem deshalb beschrieben, weil sein Lehrer fest davon überzeugt war, dass es sich um eine eigene Art handelte. Bei Sauter ist daher zu lesen: "Dahlica seileri (Sauter). Diese Art wurde nach Tieren von Goppenstein VS beschrieben (Sauter, 1954). Sie wurde auch zwischen Altanca und Brugniasco im Tessin gefunden. Die Beschreibung erfolgte auf Grund einer grösseren Zahl tetraploid parthenogenetischer Weibchen und eines einzigen aus einer dieser Kulturen Seilers geschlüpften Männchens, wobei die feste Überzeugung Seilers, dass es sich hier um eine von D. triquetrella verschiedene Art handle, ausschlaggebend war. Ich bin heute der Ansicht, dass es sich doch nur um eine Form von D. triquetrella handeln dürfte. Die Unterschiede in der Sackform scheinen mir zu wenig beweiskräftig, und die abweichende Schuppenform des einzigen Männchens liesse sich meiner Meinung nach erklären, wenn es sich bei diesem Männchen um ein tetraploides Tier gehandelt hätte. Seiler hat damals diese Möglichkeit ausgeschlossen. Eine zytologische Untersuchung dieses Männchens liegt leider nicht vor."
Bei Sauter & Hättenschwiler (1991) wurde das Taxon wieder als Art gelistet, allerdings ist in der Fußnote zu lesen: "Dahlica seileri ist möglicherweise nur eine tetraploide Form von Dahlica triquetrella." Hättenschwiler (1997) lehnt die Artberechtigung vollständig ab und zitiert dazu nur die Zeilen von Sauter (1983). SwissLepTeam (2010) erwähnen das Taxon auch nicht in einer der vielen Anmerkungen. Arnscheid & Weidlich (2017) führen das Taxon jetzt als Notlösung provisorisch als Unterart von D. triquetrella. Genetisch untersucht wurde es noch immer nicht.
(Autor: Erwin Rennwald)
4.6. Literatur
- Arnscheid, W.R. & M. Weidlich (2017): Psychidae. — In: Karsholt, O., Mutanen, M. & M. Nuss [edit.]: Microlepidoptera of Europe. Volume 8: 1-423; Leiden & Boston (Brill).
- Dahl, F. (1911): Wieder eine flohähnliche Fliege. — Zoologischer Anzeiger 28 (9/10): 212-221 [Digitalisat auf archive.org].
- Erstbeschreibung der Gattung Dahlica: Enderlein, G. (1912): Zur Kenntnis der Zygophthalmen. Über die Gruppierung der Sciariden und Scatopsiden. — Zoologischer Anzeiger 40 (10/11): 261-282 [Digitalisat auf archive.org].
- Hättenschwiler, P. (1997): Dahlica triquetrella. — In: Pro Natura – Schweizerischer Bund für Naturschutz (Hrsg.) (1997): Schmetterlinge und ihre Lebensräume. Arten, Gefährdung, Schutz. Schweiz und angrenzende Gebiete. Band 2: 185-187. Egg (Fotorotar AG).
- Erstbeschreibung: Hübner, J. [1796-1834]: Sammlung europäischer Schmetterlinge 8: pl. 1-71.
- Beschreibung als Solenobia seileri: Sauter, W. (1954): Zur Morphologie und Systematik der schweizerischen Solenobia-Arten (Lep. Psychidae) (Vorläufige Mitteilung). — Mitteilungen der Schweizerischen Entomologischen Gesellschaft, Bulletin de la Société Entomologique Suisse, 27 (4): 429-434.
- Sauter, W. & P. Hättenschwiler (1991): Zum System der palaearktischen Psychiden (Lep., Psychidae). 1. Teil. Liste der palaearktischen Arten. — Nota lepidopterologica 14 (1): 69-89. [PDF auf zobodat.at]
- Sauter & Hättenschwiler (2004): Zum System der palaearktischen Psychidae. 3. Teil: Bestimmungsschlüssel für die Säcke. — Nota lepidopterologica 27 (1): 59–69 [Digitalisat auf archive.org].
- SCHÜTZE (1931): 12
- Sobczyk, T. (2011): Psychidae. World Catalogue of Insects 10. — 467 S.; Stenstrup (Apollo Books)
- Štys, P. (1960): On the lepidopterous nature of the previously dipterous genus Dahlica Enderlein, 1912 (Lepidoptera, Psychidae – Diptera, Fungivoroidea). — Časopis Československé společnosti entomologické 57 (1): 76-83.
- SwissLepTeam (2010). Die Schmetterlinge (Lepidoptera) der Schweiz: Eine kommentierte, systematisch-faunistische Liste. — Fauna Helvetica 25. Neuchâtel (CSCF & SEG).
- Weidlich, M. (2014): Zum Vorkommen der bisexuellen Form von Dahlica triquetrella (Hübner, 1813) in Europa, insbesondere in Ungarn und in der Slowakei (Lepidoptera: Psychidae). — Microlepidoptera.hu 7: 61–66. [PDF-Version auf epa.oszk.hu]
- Weidlich, M. (2015): Beobachtungen zur Psychidenfauna Italiens mit der Neubeschreibung einer Dahlica Enderlein, 1912 (Lepidoptera: Psychidae). — Linzer biologische Beiträge 47 (2): 1909-1934 [PDF auf zobodat.at].
