

 +14Kontinente:EU
+14Kontinente:EU













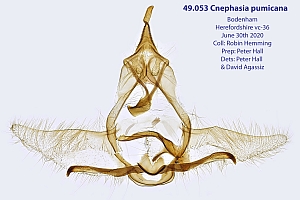


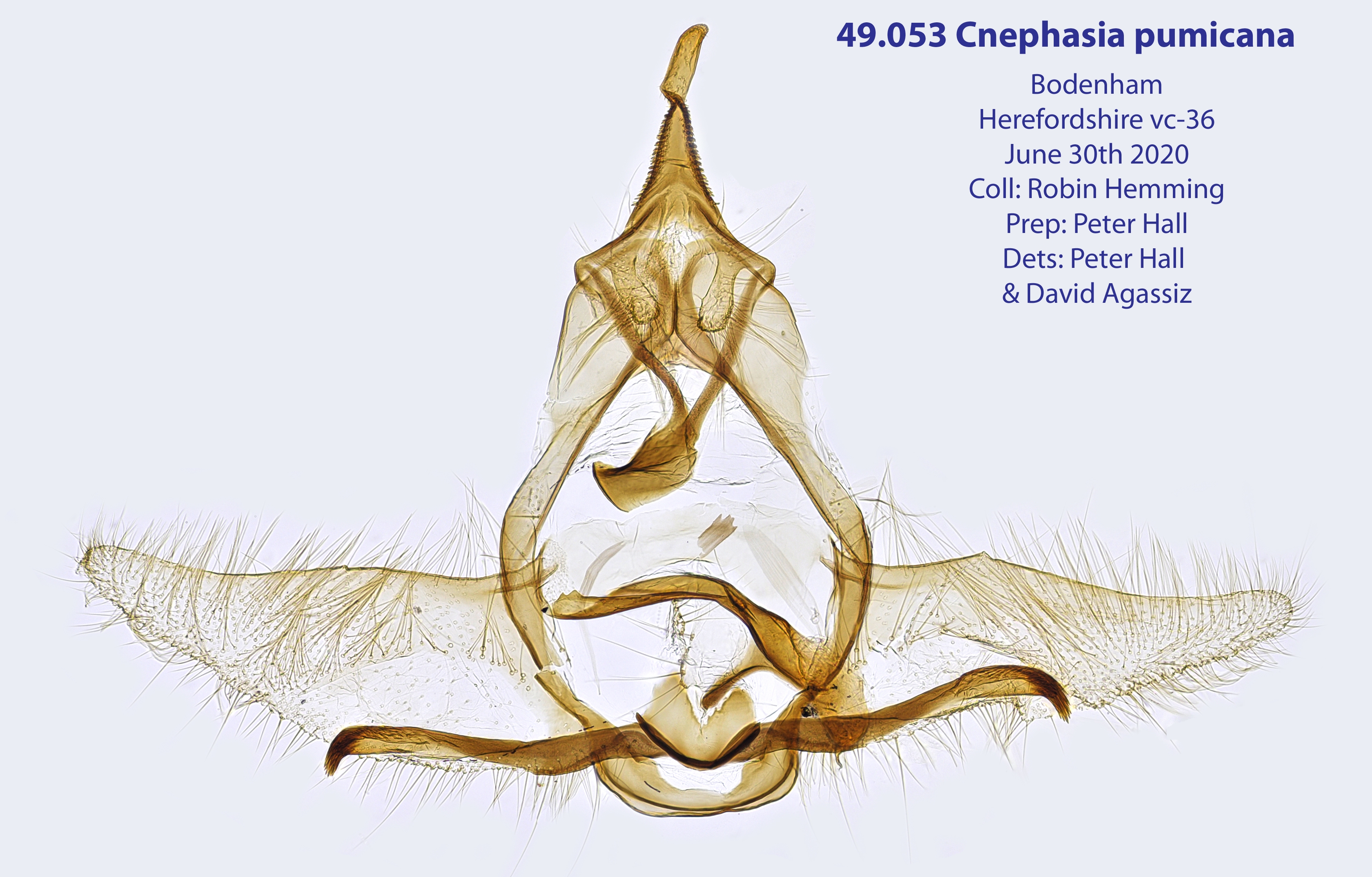
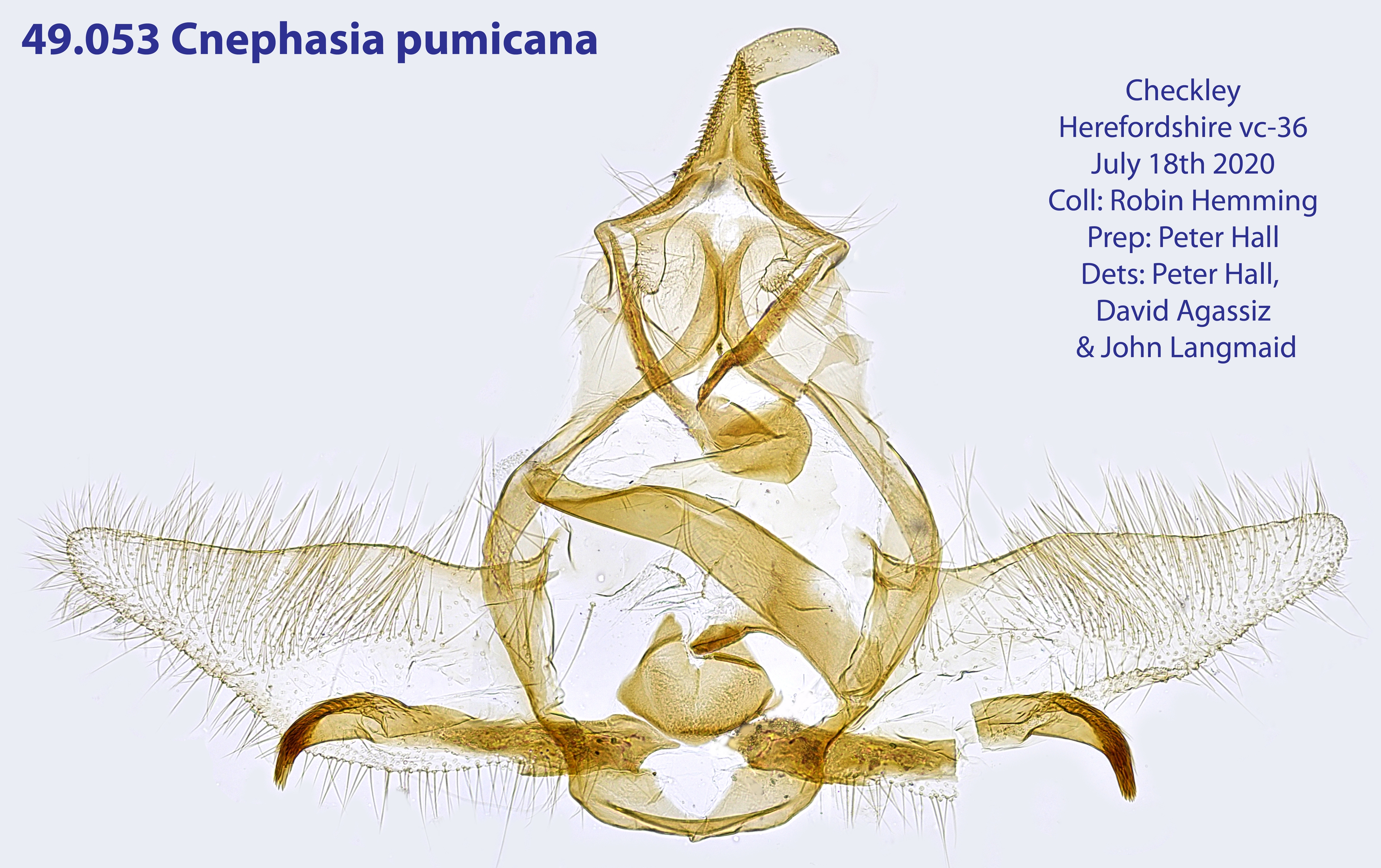
2. Diagnose
2.1. Genitalien
Vergleiche mit Cnephasia pasiuana, Cnephasia genitalana und weiteren verwechslungsträchtigen Arten finden sich auf [Mothdissection].
2.1.1. Männchen
2.2. Erstbeschreibung


3. Biologie
3.1. Nahrung der Raupe
- [Poaceae:] Avena sp. (Hafer)
- [Poaceae:] Secale sp. (Roggen)
- [Poaceae:] Triticum sp. (Weizen)
- [Poaceae:] Hordeum sp. (Gerste)
- [Poaceae:] Elymus sp. [= Agropyron sp.] (Quecke)
Hancock et al. (2015: 97-98) melden: "Ovum. Laid on trees or other suitable in late June and July." Und dann zur Raupe: "August - June. The larva overwinters in its first instar, then in early spring it leaves its hibernaculum and moves to a suitable food plant, sometimes assisted by the wind. It is highly polyphagous on cereals, grasses, legumes and flax; some 22 host plants have been recorded (Chambon & Genestier, 1980)." [in dem zitierten Artikel werden keine konkreten Pflanzenarten gelistet, wohl aber in anderen dort zitierten Artikeln der Autoren.]
Raupenfunde gelangen früher üblicherweise erst in den letzten Raupenstadien im Frühjahr, wo sich die Raupen als typische Wickler-Raupen zeigen. Die Ausführungen von [eurasian-tortricidae.linnaeus.naturalis.nl ["Meijerman, L. & S.A. Ulenberg (2000)"]] zu "Cnephasia pasiuana" gehören nicht zu jener Art, sondern zur später wieder abgetrennten C. pumicana und liefern sehr interessante Angaben zu den frühen Stadien: "Adults emerge from the middle of June to the beginning of August. The moths have a presexual period of about 1-5 days after emergence on their host plants. Copulation takes place in the early twilight inside the fields. One day after copulation the females fly to nearby wood (forest, wind shelters, solitary trees etc.), where they deposit their eggs on the rough surface of the bark until midnight. The eggs are white, but later change to orange-red. Eggs hatch after about 14 days and the neonate larvae (about 1 mm long) first crawl over the bark for a period of about 2 days. They then conceal themselves in crevices of the bark or moss and weave a small white cocoon (the hibernaculum). They stay inside the cocoon for the rest of the summer, autumn and winter in a state of dormancy, first an obligate diapause until mid to late December and later a quiescence that lasts until the beginning of the spring. The quiescence ends when the maximum daily temperature exceeds 8 °C (as the climate is in March in Central Europe). The larvae then leave their hibernacula. Migration on a massive scale starts on sunny days, when the average temperature reaches 13 °C (usually from mid to late April). The larvae move on the surface of the bark and then descend on threads on which they are carried by the wind. Larvae that land on suitable host plants start to feed on the leaves. The larvae mine in narrow oblong passages, feeding on the parenchyma tissue between the upper and lower epidermis. Late third instar larvae leave the mines and continue to feed on the leaves in the typical manner of leafrollers, by folding and spinning the leaf margins towards the central veins. Pupation commences in June and lasts 9-25 days, depending on the ambient temperature. Pupation always occurs on the host plant, mainly in the sheath of the flag leaf or between the grains and the awns (Glas, 1991)." Zu den Nahrungspflanzen der Raupe heißt es dann: "Host plants: This species is a pests and of certain economical importance on wheat (Triticum), barley (Hordeum), rye (Secale) and oat (Avena). Host plants also include Aster, Anthemis, Achillea, Artemisia, Centaurea, Chrysanthemum, Circium [recte: Cirsium], Colchicum, Erigeron, Echium, Hyochoeris, Ranunculus and Thalictrum." Und zu den Schäden ist dann noch zu erfahren: "Larvae cause yield-reducing damage to cereal crops. From the beginning of the formation of the ears, the sheath of the flag leaf is penetrated by the larvae and stamens and pistils are destroyed. In some cases, even stalks are cut. They feed in the developing grain until milky-ripeness. The causes of outbreaks of Cnephasia pasiuana seem to be climatic factors (probably dry periods during larval dispersion) and the high proportion of cereals in crop rotations. Populations may be effectively controlled by natural enemies. Economic damage is now only reported from Germany and France (Glas, 1991)." [Die Angaben zu den Nicht-Süßgräsern dürften sich wweitgehend tatsächlich auf Cnephasia pasiuana beziehen.]
(Autor: Erwin Rennwald)
4. Weitere Informationen
4.1. Andere Kombinationen
- Sciaphila pumicana Zeller, 1847 [Originalkombination]
4.2. Taxonomie
Lange Jahre galt Cnephasia pumicana als eigenständige Art. Dann wurde sie vielfach in die Synonymie von C. pasiuana eingezogen. Die Berechtigung dieses Vorgehens darf angesichts pheromonbiologischer und auch (geringer) genitalmorphologischer Unterschiede hinterfragt werden. Langmaid & Agassiz (2010) haben C. pumicana wieder zur bona species erklärt. Auch von der [Fauna Europaea] werden beide Arten wieder akzeptiert. Zur Diskussion siehe auch den [Forumsbeitrag von W. Nässig].
4.3. Faunistik
Locus typicus ist Syrakus auf Sizilien (Italien).
Geiter, Homma & Kinzelbach (2002) führen die Art als Neozoon mit dem Status C (fraglich) für Deutschland. Gaedike in Gaedike et al. (2017: 168) meldet: "Nach Langmaid & Agassiz (2010) ist pumicana (Zeller, 1847) eine valide Art und nicht ein Synonym zu C. pasiuana (Hübner, 1799). Die bisherigen Nachweise für pasiuana müssen deshalb geprüft werden. Gesicherte Meldung für BY (Haslberger in litt.)."
Huemer & Rabitsch (2002) führen die Art für Österreich ebenfalls bei den Neobiota; sie halten die Art für etabliert und für nicht expansiv. Schadmeldungen aus Österreich sind ihnen (nach Henning 1986) seit 1976 bekannt.
Nach der [Fauna Europaea] kommt die Art in Deutschland, Österreich, Tschechien, Italien, in der Slowakei sowie auf Zypern und den Dodekanesischen Inseln vor. Die Deutung als Neozoon ist schlecht begründet. Sie darf nicht an Schadmeldungen festgemacht werden. Aus Westeuropa scheint es nur die Angaben von Langmaid & Agassiz (2010) aus Großbritannien zu geben, aus der Westmediterraneis gar keine.
(Autor: Erwin Rennwald)
4.4. Literatur
- Bathon, H. (1981): Zur Erfassung und Bestimmung einiger mitteleuropäischer Cnephasia-Arten (Lep.: Tortricidae). — Entomologische Zeitschrift 91 (22): 248-252. Ex libris Jürgen Rodeland.
- Bathon, H. & M. Glas (1983): Zur Verbreitung des Getreidewicklers, Cnephasia pumicana Zeller (Lepidoptera: Tortricidae), in der Bundesrepublik Deutschland. Erste Ergebnisse einer Pheromonfallen-Erhebung von 1982. — Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 35 (6): 81-86. [PDF auf ojs.openagrar.de]
- Bengtsson, B. Å. (2023): Anmärkningsvärda fynd av småfjärilar (Microlepidoptera) i Sverige 2022. — Entomologisk Tidskrift 144 (1-2): 1-20. [Sekundärzitat, Abstract auf et-online.nu]
- Chambon, J.-P. & G. Genestier (1980): Identification et biologie des espèces françaises les plus communes du genre Cnephasia Curtis [Lep. Tortricidae]. — Bulletin de la Société entomologique de France, 85 (3-4): 71-79. [PDF auf persee.fr]
- Chambon, J.-P. (1981): Répartition géographique de Cnephasia pumicana Zeller en France [Lep. Tortricidae]. Utilisation d'une phéromone sexuelle de synthèse. — Bulletin de la Société entomologique de France, 86 (1-2): 26-29. [PDF auf persee.fr]
- Gaedike, R., Nuss, M., Steiner, A. & R. Trusch (2017): Verzeichnis der Schmetterlinge Deutschlands (Lepidoptera). 2. überarbeitete Auflage. — Entomologische Nachrichten und Berichte (Dresden), Beiheft 21: 1-362.
- Gebhardt, H., Kinzelbach, R. & S. Schmidt-Fischer (Hrsg.) (1996): Gebietsfremde Tierarten. Auswirkung auf einheimische Arten, Lebensgemeinschaften und Biotope. Situationsanalyse. — 314 S., Landsberg (ecomed verlagsgesellschaft). [Sekundärzitat nach Geiter, Homma & Kinzelbach (2002)]
- Geiter, O, Homma, S. & R. Kinzelbach (2002): Umweltforschungsplan des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit. Forschungsbericht 296 89 901/01. UBA-FB 000215: Bestandsaufnahme und Bewertung von Neozoen in Deutschland Untersuchung der Wirkung von Biologie und Genetik ausgewählter Neozoen auf Ökosysteme und Vergleich mit den potenziellen Effekten gentechnisch veränderter Organismen. I-III, 1-74, Anhang 1 1-35, Anhang II 1-31, Anhang III 1-52 [PDF auf umweltbundesamt.de].
- Glas, M. (1985): Zweiter Beitrag zur Verbreitung von Ährenwickler, Cnephasia longana (Haworth) und Getreidewickler, C. pumicana (Zeller), (Lepidoptera, Tortricidae) in der Bundesrepublik Deutschland. Ergebnis einer Pheromonfallenerhebung von 1983. — Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 37: 21–27.
- Glas, M. (1986): Zur Biologie, Ökologie, Verbreitung und Bekämpfung von Wicklern an Getreide, Cnephasia longana (Haworth) und C. pumicana (Zeller). – Hannover. [Sekundärzitat]
- Hancock, E.F., Bland, K.P. & J. Razowski (2015): The moths and butterflies of Great Britain and Ireland. Volume 5 (Part 1). Tortricidae, Tortricinae & Chlidanotinae. - 245 S.; Leiden & Boston (Brill).
- Henning, H. (1986): Der Getreidewickler, Cnephasia pumicana (Zeller), und der Ährenwickler, Cnephasia longana (Haworth) (Lepidoptera: Tortricidae) in Österreich. — Pflanzenschutz 2 :8-10. [Sekundärzitat]
- Huemer, P. & W. Rabitsch (2002): 6.3.19 Schmetterlinge (Lepidoptera). - 354-362. In: Essl, F. & W. Rabitsch (2002): Neobiota in Österreich. - 432 S.; Umweltbundesamt, Wien.
- Langmaid, J. R. & D. J. L. Agassiz (2010): Cnephasia pumicana (Zeller, 1847) (Lep.: Tortricidae) stat. rev. newly recognised as British. — The Entomologist's Record and Journal of Variation 122 (4): 137-142.
- Erstbeschreibung: Zeller, P. C. (1847): Bemerkungen über die auf einer Reise nach Italien und Sicilien beobachteten Schmetterlingsarten. — Isis von Oken 1847 (2): 121-159, (3) 213-233, (4) 284-308, (6) 401-457, (7) 481-522, (8) 561-594, (9) 641-673, (10) 721-771, (11) 801-859, (12) 881-914.
4.5. Informationen auf anderen Websites (externe Links)
- [inra.fr]
- [hantsmoths]
- [mothdissection.co.uk] (auch mit Lebendbild)
